To set up the lab, we attached three electrode tabs to the subject; a black tab (ground) was placed on the calf area, while both the green (positive) and red (negative) electrode tabs were attached to the quadriceps, with the red tab more proximal to the hips than the green tab.


The first test administered measured a voluntary response; the test subject sat on a desk with their eyes closed, and a hammer was struck against the desk, generating a loud sound. After hearing this sound, the test subject's leg rapidly lifted into the air. The nerve activity was measured on the computer system, and the time between the hammer striking the desk and the reaction itself was measured to demonstrate a specific reaction time of .23 seconds.

During the second test, the involuntary reflexes of the patellar tendon were measured. To collect this data, the test subject became relaxed, again with closed eyes, and was tapped with a small mallet on a contact point of the tendon. The nerve activity of the patellar tendon in the quadriceps was measured, as well as the time that lapsed between the initial contact and the reflex that occurred, .07 seconds.

The third test administered measured not only reflexes, but also reinforced reflexes. For this test, the subject sat with eyes closed, as in the other tests, but also hooked their hands together at chest level (like one would in a traditional choir). As in Test 2, the patellar tendon was struck with a small mallet, however, every time a reflex occurred, the subject was asked to pull their hands away without actually breaking the connection between their hands. This movement reinforced the reflex that was visible in the legs. Again, both nerve activity and time between the stimulus and reaction were recorded, this time being .04 seconds.

Test 4 was a repetition of Test 1, again measuring voluntary reaction, however, this test used a larger hammer to generate the sound. The reaction time in this test was equal to .22 seconds.

Test 5 was also a repetition of Test 2, the difference being the use of a larger mallet to administer the strike to the patellar tendon. The reaction time from this test was .04 seconds.
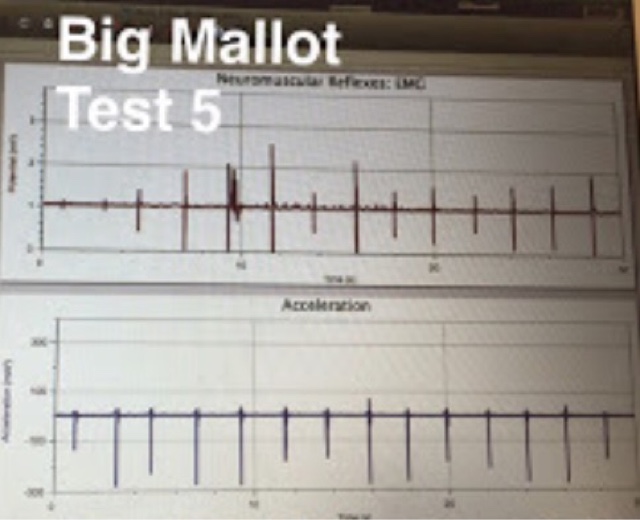
The same procedure was followed for Test 6 as was followed during Test 3, the difference being the size of the mallet used during the test. For Test 6, a larger mallet was used to strike the patellar tendon than the smaller mallet used during Test 3. The reaction time for this test was .04 seconds.

To visualize and more closely compare the reaction times as well as the nerve activity, the data from each test was organized into the following graphs:
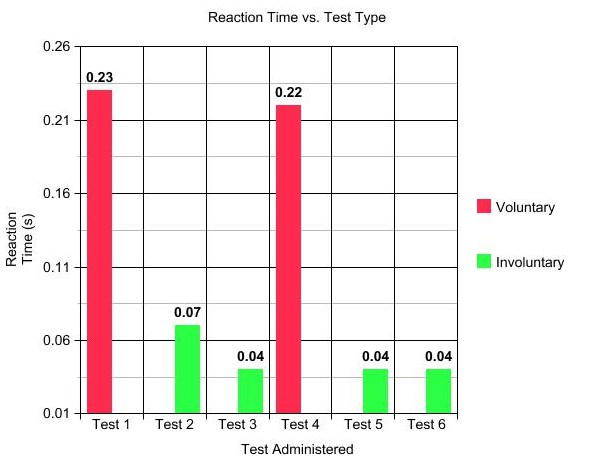
It is very likely that the reaction time was slower during the voluntary tests due to the fact that there was no direct stimulus to the nerves, only voluntary instinct. The subject had to process the sound and think about moving their leg before any type of reaction ever occurred, while the involuntary tests interacted with the nerves directly, forcing the subject to move before they even had a chance to voluntarily process the idea that a stimulus had occurred.
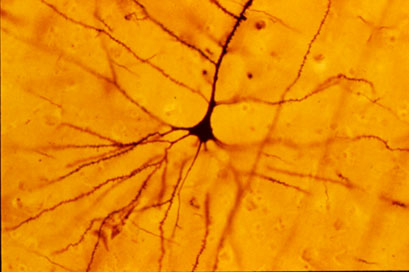
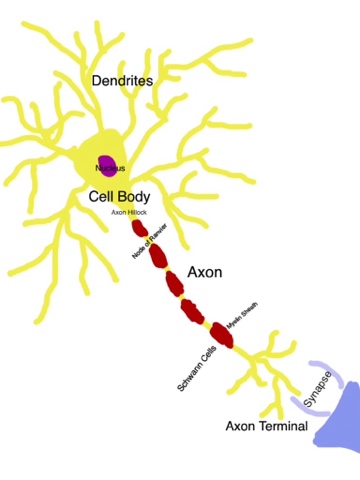
It was very interesting to visually picture the electrical activity of nerves, as well as analyze why reaction times differ depending on the type of reaction caused through nerve stimulus.



























{kind=link}